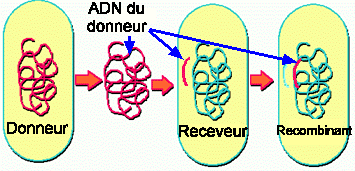
Figure 1. Schéma général de l'échange génétique bactérien. L'ADN du donneur est transféré au receveur où il remplace un ou plusieurs gènes par recombinaison.
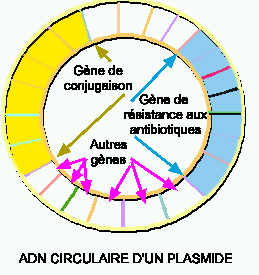
Figure 2. LE PLASMIDE FACTEUR DE FERTILITÉ OU FACTEUR F. Le facteurF porte les gènes pour la conjugaison ainsi que d'autres gènes tels que des séquences d'insertion, qui permettent l'intégration du plasmide dans le chromosome de la cellule hôte, et des gènes de résistance aux antibiotiques.
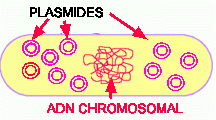
Figure 3. Des plasmides dans une bactérie hôte. Une cellule peut contenir 1 plasmide, plusieurs copies d'un plasmide et/ou plusieurs plasmides différents.
Les plasmides sont de tailles variées, entre 1000 et 25000 paires de bases vs 4 millions pour le génome. Les plasmides se répliquent de façon autonome par rapport au chromosome. Une cellule hôte peut contenir un plasmide, plusieurs copies d'un plasmide et/ou plusieurs plasmides différents. Les plasmides portent généralement des gènes qui ne sont pas essentiels pour la survie de la cellule, sauf dans des conditions exceptionnelles. Par exemple, plusieurs plasmides portent des gènes de résistance aux antibiotiques. Lorsque le plasmide de résistance donné est présent dans la cellule, elle ne sera pas affectée par l'antibiotique correspondant. Mais si ce plasmide est perdu, la cellule devient vulnérable à l'antibiotique. Certains plasmides portent des gènes de résistance à plusieurs antibiotiques, ce qui en fait de dangereux pathogènes. Des maladies telles la diphtérie et le botulisme sont dues à des plasmides de virulence qui portent des gènes exprimant des toxines. Ce moyen permet en fait à ces bactéries pathogènes de mieux résister au système immunitaire de leur hôte. Un cas récent de bactérie portant un plasmide de virulence est la souche de E.coli O157:H7 causant des colliques hémorragiques. D'autres plasmides portent des gènes protecteurs contre des susbtances toxiques telles le mercure, le cuivre, etc et enfin d'autres permettent l'utilisation de substrat exotiques comme des hydrocarbones aromatiques, des pesticides, et des sucres.
L'existence des plasmides permet donc à un ensemble de cellules de conserver un patrimoine génétique qui peut être très utile dans des conditions particulières, mais qui viendrait cependant alourdir le génome de ces cellules si chacune d'entre elle devait porter tous ces gènes. Dans ce dernier cas, la dépense énergétique qui s'ensuivrait serait fort probablement trop importante pour le bénéfice acquis, ce qui rendrait le micro-organisme non-viable (ou plus précisément, non-concurrentiel!). Cependant, dans le cas où un plasmide serait requis, dû à des conditions environnementales difficiles, l'environnement exercerait une PRESSION SÉLECTIVE favorisant la survie des cellules portant ce plasmide, les seules capables de survivre et de se multiplier.
D'un point de vue technologique, on peut exploiter la capacité des plasmides à transférer du matériel génétique, pour introduire des gènes d'intérêt dans des souches cellulaires ( Chap X).
![]() Lyser
la membrane cellulaire du donneur et isoler son ADN de manière à
obtenir une solution pure, sans autres composants cellulaires (ADN NU)
Lyser
la membrane cellulaire du donneur et isoler son ADN de manière à
obtenir une solution pure, sans autres composants cellulaires (ADN NU)
![]() Ajouter
l'ADN nu à la cellule receveuse dite COMPÉTENTE,
c'est à dire capable de lier l'ADN et l'introduire dans son cytoplasme.
La fréquence de transformation chez des cellules très compétentes
se situe aux environs de E-3 pour la plupart des espèces lorsqu'il
y a un excès d'ADN.
Ajouter
l'ADN nu à la cellule receveuse dite COMPÉTENTE,
c'est à dire capable de lier l'ADN et l'introduire dans son cytoplasme.
La fréquence de transformation chez des cellules très compétentes
se situe aux environs de E-3 pour la plupart des espèces lorsqu'il
y a un excès d'ADN.
![]() L'ADN
nu pénètre dans la cellule receveuse où il peut être
échangé avec l'ADN de la cellule ou, si c'est un plasmide,
se répliquer.
L'ADN
nu pénètre dans la cellule receveuse où il peut être
échangé avec l'ADN de la cellule ou, si c'est un plasmide,
se répliquer.
La transformation peut être utilisée pour échanger des gènes entre des bactéries, des plantes et des animaux. Dans chaque cas, la méthode utilisée diffère sur les détails. Chez les bactéries, la "compétence" ne peut pas être prédite a priori pour une souche donnée. Cependant, le traitement suivant induit généralement la compétence chez les gram négatives:
![]() Des
cellules jeunes sont incubées dans une solution de chlorure de calcium
pour approx. 30 min. sur de la glace. Dans certains cas, un sel de magnésium
est aussi utilisé.
Des
cellules jeunes sont incubées dans une solution de chlorure de calcium
pour approx. 30 min. sur de la glace. Dans certains cas, un sel de magnésium
est aussi utilisé.
![]() Les
cellules sont concentrées et mise en suspension dense dans la solution
de calcium.
Les
cellules sont concentrées et mise en suspension dense dans la solution
de calcium.
![]() De
l'ADN nu est alors mélangé avec la suspension de cellules
compétentes sur de la glace pour approx. un autre 30 min., suivi
d'un léger chauffage
De
l'ADN nu est alors mélangé avec la suspension de cellules
compétentes sur de la glace pour approx. un autre 30 min., suivi
d'un léger chauffage
![]() Les
cellules transformées sont incubées dans un milieu riche
pour approx. 1 à 1.5 h, puis mises en plat sur du milieu contenant
un agent sélectif.
Les
cellules transformées sont incubées dans un milieu riche
pour approx. 1 à 1.5 h, puis mises en plat sur du milieu contenant
un agent sélectif.
Plusieurs
autres
techniques de transformation existent pour les cellules eucaryotes,
dont les mélanges de certains sels avec
l'ADN. Ces sels se lient à l'ADN et aident à sa pénétration
dans la cellule. Les cellules de plantes sont souvent recouvertes d'une
épaisse paroi, difficile à pénétrer. Pour les
transformer, on enduit des microbilles de platine avec l'ADN du donneur
et on projette ces particules sur les cellules à l'aide d'un "pistolet
à ADN". On peut aussi soumettre les cellules receveuses
à des différences de potentiel électrique important
qui vont rendre la membrane plus perméable à l'ADN (ÉLECTROPORATION).
Étant donné la nature universelle de l'ADN, des transferts
de matériel génétique sont possibles entre les espèces.
Des animaux ou des plantes qui ont été transformés
par de l'ADN d'une autre espèce sont dits TRANSGÉNIQUES.
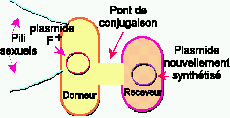
Figure 4. Conjugaison bactérienne.
La découverte d'une forme d'échange entre les bactéries, la CONJUGAISON, dans les années 50 a étonné autant les scientifiques que les non-spécialistes. Sa similitude avec le mode de reproduction humain était aussi amusante pour certains que choquante pour d'autres. Depuis sa découverte, on a montré que l'échange d'ADN par conjugaison était plus fréquente qu'on ne le pensait au début. Initialement, on croyait que la conjugaison ne pouvait se produire qu'entre membres de la même espèce, ou d'espèces rapprochées. Cependant, l'accumulation des preuves a démontré que la conjugaison se produisait aussi entre différentes espèces de bactéries et même entre des bactéries et certains eucaryotes.
Les conditions de base pour la conjugaison sont:
![]() Les
cellules donneuses doivent contenir un plasmide
de fertilité appelé facteur
F, un plasmide portant les gènes permettant la conjugaison.
Les cellules contenant le facteur F sont désignées F+
ou MALE alors que les cellules sans
facteur F sont désignées F-
ou FEMELLE.
Les
cellules donneuses doivent contenir un plasmide
de fertilité appelé facteur
F, un plasmide portant les gènes permettant la conjugaison.
Les cellules contenant le facteur F sont désignées F+
ou MALE alors que les cellules sans
facteur F sont désignées F-
ou FEMELLE.
![]() Le facteur
F est responsable de la synthèse des pili sexuels. Ces pili sont
de long tubes minces de protéines, pourvus à leur extrémité
d'un récepteur ayant une affinité pour des molécules
(ligands) à la surface des cellules receveuses.
Le facteur
F est responsable de la synthèse des pili sexuels. Ces pili sont
de long tubes minces de protéines, pourvus à leur extrémité
d'un récepteur ayant une affinité pour des molécules
(ligands) à la surface des cellules receveuses.
![]() À la
suite de l'adhésion de deux cellules par l'intermédiaire
des pili, ils s'unissent via un "CANAL DE TRANSFERT".
La nature de cette union n'est pas encore claire, non plus que le rôle
exact des pili sexuels dans sa formation. Cependant, ce qui est reconnu,
c'est qu'il y a un conduit d'ouvert par lequel l'ADN peut passer de la
cellule donneuse à la cellule receveuse.
À la
suite de l'adhésion de deux cellules par l'intermédiaire
des pili, ils s'unissent via un "CANAL DE TRANSFERT".
La nature de cette union n'est pas encore claire, non plus que le rôle
exact des pili sexuels dans sa formation. Cependant, ce qui est reconnu,
c'est qu'il y a un conduit d'ouvert par lequel l'ADN peut passer de la
cellule donneuse à la cellule receveuse.
![]() Une enzyme
spécial COUPE un des brins de
l'ADN du donneur à un endroit précis et un brin
fraichement synthétisé d'ADN passe par le canal
de transfert dans la cellule receveuse.
Une enzyme
spécial COUPE un des brins de
l'ADN du donneur à un endroit précis et un brin
fraichement synthétisé d'ADN passe par le canal
de transfert dans la cellule receveuse.
![]() Ce nouveau
brin est converti en double brin qui est alors capable d'échange
avec le génome de l'hôte par recombinaison.
Ce nouveau
brin est converti en double brin qui est alors capable d'échange
avec le génome de l'hôte par recombinaison.
La forme la plus fréquente de conjugaison implique
le transfert de plasmides d'une cellule
à une autre. Ce processus est très efficace. Par exemple
la cellule receveuse qui recoit le facteur F devient elle-même F+
et commence aussitôt à conjuguer avec ces voisines. Certains
plasmides de fertilité vont fusionner avec l'ADN
génomique de la cellule hôte et ainsi transférer
tout le génome de la cellule lors d'une prochaine conjugaison. Dans
ces conditions, n'importe lequel des gènes du donneur peut être
introduit dans le génome d'une cellule receveuse.
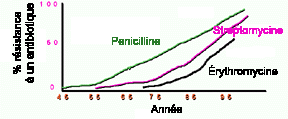

Figure 5. L'augmentation des bactéries résistantes à 3 antibiotiques en fonction du temps et un schéma d'une superbactérie avec plusieurs gènes de résistance.
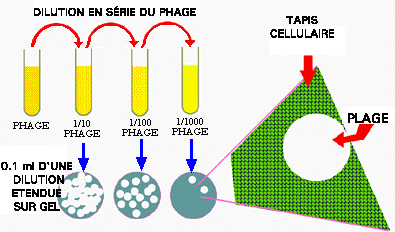
Figure 6. Technique de compte viral: le compte de plage de lyse
D'une autre façon, l'addition de phage dans une culture bactérienne en milieu liquide causera la disparition des bactéries en quelques minutes. On a déjà pensé utiliser les phages comme agents thérapeutiques contre des infections bactériennes. Bien que ces essais n'aient pas fonctionné, à cause de la réponse immunitaire de l'organisme contre les phages, on a découvert que les phages étaient extrêmement spécifiques quant aux choix de leur hôte. En fait, cette spécificité était tellement grande que les phages ont été utilisés par la suite pour identifier certaines souches bactériennes.
La capacité du phage de transporter de l'ADN bactérien
a été découverte en 1952. Des scientifiques ont alors
découvert qu'en séparant deux bactéries génétiquement
différentes par une membrane de très faible taille de pores,
ils pouvaient encore observer des transferts génétiques entre
elles. Il ne s'agissait pas de transformation puisque le transfert ne pouvait
pas être empêché par l'utilisation de la DNase. Ils
ont alors supposé que l'ADN devait être protégé
de la DNase par quelque chose. Ce quelque chose, c'était en fait
un bactériophage infectant une des souches. Les phages sont composés
de matériel génétique entouré de protéines
qui, en effet, protègent l'ADN contre la DNase. Ce processus de
transfert par l'intermédiaire de phages est appelé TRANSDUCTION.
![]()
Figure 7. La transduction.
La transduction fonctionne comme suit.Un phage infecte une bactérie et injecte sont ADN dans l'hôte. L'ADN viral prend le contrôle du métabolisme de l'hôte, le transformant en "usine à phage". Il dirige la production des composants du phage incluant l'ADN viral. Lors d'une des dernières étapes du cycle "vivant" du phage les différentes composantes du phage sont assemblées en phages complets et la cellule est rompue pour relâcher les nouvelles particules. Durant le processus d'assemblage, des erreurs très rares peuvent se produire, faisant en sorte que des fragments d'ADN de l'hôte soient introduits dans le nouveau phase. Puisque qu'il n'y a pas de place dans la capside pour tout ce matériel génétique, dans ces rares cas, il doit manquer des gènes viraux, ce qui créé des phages défectifs, incapable de se multiplier. Cependant, ces phages défectifs ont toujours la capacité de sortir de la cellule, de s'attacher à un nouvel hôte et d'introduire le gène qu'il porte dans la nouvelle cellule. Puisque le phage défectif ne porte que de l'ADN bactérien, l'infection de l'hôte ne se produit pas. Par contre, une recombinaison entre l'ADN de l'hôte précédent et le nouvel hôte, et les allèles transportées par le phage dans ce nouvel hôte peuvent être incorporées dans l'ADN de l'hôte, se faisant changeant son phénotype (Fig. 7). Pour détecter ce processus, les nouveaux hôtes sont cultivés sur un milieu qui permet de démontrer la présence d'une allèle particuliaire provenant de l'hôte précédent.
Par exemple, si les phages de transduction sont tout d'abord cultivés sur des hôtes bactériens portant un gène de résistance à la pénicilline, un nombre très faible de phages vont incorporer le gène de résistance à la pénicilline. Si le phage de cette cuture est subséquemment utilisé pour infecter des hôtes sensibles à la pénicilline et que les survivants sont mis en plat sur un milieu contenant de la pénicilline, seulement ces rares bactéries ayant recu à la fois le gène de résistance à la pénicilline et un gène défectif de phage et l'on inséré dans leur propre génome vont produire des colonies sur le milieu.
QUESTION: "Si l'évenement est si rare, comment se fait-il que nous puissions l'observer?".
RÉPONSE: La réponse est dans le nombre prodigieux de phage et de bactéries avec lesquels l'on peut travailler. Un stock de phage peut contenir 1E11 phage/ml et une culture bactérienne 2E9 bactéries/ml. Donc, même une probabilité de 1E-8 a de bonnes chances de se réaliser.
Puisque les phage sont faciles à cultiver et qu'une fois obtenus on peut les stocker et les utiliser plus tard, la transduction est un moyen pratique et efficace pour générer toutes sortes de gènes d'une bactérie et en faire l'étude.